Dossier
LES GISEMENTS DE VERTEBRES FOSSILES DU TARN
HISTORIQUE DES DECOUVERTES ET REVUE BIBLIOGRAPHIQUE
L'OLIGOCENE
PALEOCLIMATS ET PALEOENVIRONNEMENTS
Le début de l’Oligocène est marqué par une nouvelle crise globale, « The Terminal Eocene Event » (TEE). Cet évènement, lié principalement à la tectonique des plaques, est marqué par une baisse générale du niveau des océans et par un important changement climatique avec diminution de la température, aridité croissante, forte accentuation du contraste saisonnier qui voit le climat européen passer d’un type tropical à un type subtropical, voire tempéré ou méditerranéen. Les forêts tropicales se cantonnent dorénavant aux régions équatoriales et l’Antarctique se couvre d’une calotte glaciaire.
Des habitats ouverts de type savane se développent aux dépens des milieux forestiers. Les saisons étant de plus en plus marquées, nos latitudes se peuplent d'arbres à feuilles caduques. Les graminées s'étendent depuis les berges des rivières. Les Angiospermes continuent leur expansion.

Les environnements aquitains sont marqués par les inondations qui ensablaient et envasaient de vastes plaines d’épandages fluviatiles sur lesquelles se juxtaposaient des forêts denses et diversifiées, des parcelles plus ouvertes à arbustes et buissons, des zones marécageuses de type palustre ou lacustre.
La fin de l’Oligocène est marquée par une nouvelle crise climatique « The Terminal Oligocene Crisis » caractérisée par une baisse brutale de la température et de l’humidité.
CARACTERISTIQUES DES FAUNES
Les modifications climatiques du début de l’Oligocène affectent directement les faunes, particulièrement dans l'hémisphère Nord. L'Europe connait une extinction massive à la fin de l’Eocène et un flux d'animaux d'origine asiatique.
Le détroit de Turgaï qui séparait l’Europe de l’Asie va partiellement s’assécher et, ne constituant plus un obstacle aux échanges fauniques, va autoriser la diffusion en Europe de mammifères d’origine asiatique aux « caractères modernes » tels les rhinocéros, les ruminants, les suidés, hamsters, écureuils, lapins, anthracothères, chalicothères… Cette vague migratoire va rapidement remplacer les mammifères endémiques européens.
Ce bouleversement dans la composition des faunes européennes à la limite Eocène - Oligocène est classiquement désigné sous le terme de « Grande Coupure », expression utilisée pour la première fois au début du XXe siècle par le paléontologue suisse Hans Georg STEHLIN (1909).
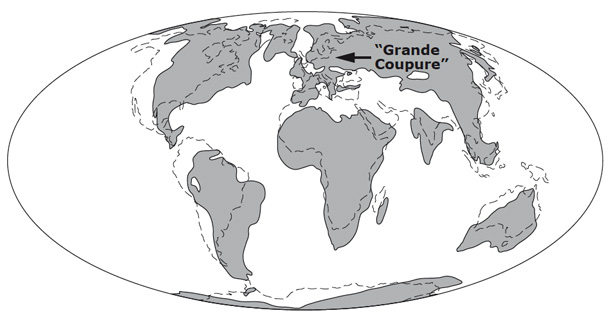
Carte paléogéographique des terres émergées à l'Oligocène (d'après Scotese, 1997)
Malgré le bouleversement des faunes représenté par « l’invasion asiatique de la Grande Coupure », les communautés mammaliennes demeurent plutôt stables, durant l’Oligocène, qui est qualifié « d’époque tranquille » par le paléontologue Stephen Jay Gould. Les changements de faune se font graduellement et la résolution stratigraphique des espèces est moins bonne qu’à l’Eocène, nécessitant d’avoir systématiquement recours à des associations d'espèces pour donner une datation.
COMPOSITION DES FAUNES
Les Artiodactyles
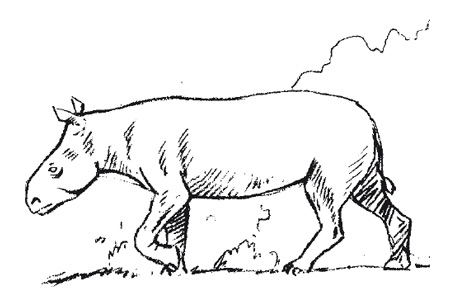
De nouvelles formes dominent les communautés mammalogiques dès la base de l’Oligocène. Parmi elles, Entelodon magnum, est qualifié par Stehlin (1909) de « chef de file de l’immigration Oligocène ». Il est présent dès la Biozone MP 21 (Gaillac).

Reconstitution de Entelodon magnum
Les Anthracothèriidés présentaient l’allure d’Hippopotames actuels et étaient vraisemblablement inféodés aux milieux palustres, avec un mode de vie quasi-amphibien supposé. Ils apparaissent dès la base de l’Oligocène S’y succèdent Anthracotherium alsaticum (MP 22) et Anthracotherium magnum que l’on retrouve dans de nombreux gisements et qui semble montrer une plus importante longévité (de MP 25 à 29).
Parmi les autres formes nous citerons Elomeryx porcinus, présent dans la Molasse de Briatexte autour des biozones MP 21 et 22, Lophiomeryx chalaniati fréquemment cité par les auteurs et dont l’extension s’étend dans le Tarn de MP 25 à MP 28, Metriotherium mirabile, de la Biozone MP 24-25 (Rabastens) à MP 27 (Tauriac). Bachiterium curtum est bien représentatif de la Biozone MP 25 (Montclar-de-Quercy)
Le genre Dremotherium apparaît à la fin de l’Oligocène, Biozones MP 28 à MP 30. Dremotherium guthi et à D. quercyi, sont tous deux présents dans le Chattien terminal de Réal-Poquemaure (Biozone MP 30). L’âge des « Dremotherium nanum et Dremotherium tolosanum « autour de MP 25 de Rabastens est possiblement à réévaluer.

Reconstitution de Anthracotherium
Les Périssodactyles
Les Périssodactyles, qui avaient dominé l’Eocène, vont connaitre un profond renouvellement avec l’arrivée des Rhinoceros, particulièrement représentatifs de l’immigration asiatique.
Parli les genres qui avaient dominé l'Eocène supérieur, le genre Plagiolophus est le seul à traverser la Grande Coupure sans trop de dommage, avec, notamment P. minus et P. fraasi qui est présent dans le Tarn jusqu’à la Biozone MP 25 (Calcaire de Biatexte). Un dernier représentant des Palaeotherium, le P. medium, atteint l'Oligocène mais ne dépasse pas la Biozone MP 21.
Dès l’Oligocène inférieur, les Rhinoceros sont représentées par trois familles : Les Hyracodontidae, Rhinocerotidae et les Amynodontidés.
- Les Hyracodontidae ne sont connus en Europe que par le genre Eggyssodon, rarement signalé dans le Tarn avec Eggysodon gaudryi à Puylaurens, autour de la Biozone MP 24-25.
- Les Rhinocerotidés sont surtout représentés par les Ronzothères qui apparaissent les formes les plus caractéristiques du début de l’Oligocène.
Ronzotherium filholi y est abondant dès les Biozones MP 21-22 dans le Tarn (Gaillac, Montans) et des formes affines sont signalées par les auteurs jusque dans MP 25, voire MP 28. Cet animal de plus d’une tonne avait une allure très gracile, ce qui lui permettait de courir avec aisance, un peu comme un cheval actuel. Contrairement aux rhinocéros actuels, il ne possédait pas de corne. Il était probablement folivore et avait une préférence pour les milieux forestiers ouverts tels que les savanes arborées.
Ronzotherium romani présent dans le Calcaire de Briatexte semble lui succéder à partir de la Biozone MP 25.
Reconstitution de Ronzotherium filholi (d'après Becker et Rauber, 2007)
Molassitherium albigense, dont l’espèce-type provient des environs de la Sauzière, près de Salvagnac, est l’espèce la plus emblématique du Chattien tarnais. Son extension semble large dans le Tarn, de la Biozone MP 24 à la Biozone MP 26 (son existence est attestée de MP 23 à MP 29 par Becker et al., 2012).
Molassitherium delemontense est signalé dans la Biozone MP 25 de Montclar-de-Quercy.
- Les Amynodontidés fournissent de bons repères biostratigraphiques dans le Tarn avec Cadurcotherium cayluxi présent dans la seule Biozone MP 25 (Puylaurens, Calcaire de Briatexte) où il est parfois associé à C. nouleti.

Reconstitution de Molassitherium albigense
Les Créodontes sont plusieurs fois signalés dans le Tarn. Le type de Hyaenodon brachyrhynchus provient des molasses stampiennes de Rabastens. Pour Muratet et al. (1992), l’âge de ce fossile serait à situer entre les Biozones MP 21 et 23. Curieusement, il est considéré par les auteurs (Biochrom’97) comme représentatif de la Biozone MP 17 du Ludien.
Hyaenodon leptorhynchus est identifié dans la Biozone MP 25 du Calcaire de Briatexte et Hyaenodon exiguus dans les Biozones MP 22 (Montclar) à MP 26-27 du Chattien des environs de Rabastens.
Les Rongeurs sont rarement mis en évidence dans les dépôts gréso-conglomératiques du Tarn. Plusieurs gisements ont cependant fourni des dents de nombreuses espèces ; Pseudoltinomys major (Montclar, MP 22) ; Issiodoromys minor, Theridomys gregarius, Blainvillimys blanvillei présents de MP 25 à MP 26, Issiodoromys pauffiensis Gliravus bruijni et Eomys zitteli plus particulièrement cantonnés à la Biozone MP 26 (Puicelci).
Les insectivores sont rarement cités, Srinitium marteli est présent à Délazens (MP 24-25), Neurogymnurus cayluxi à Montclar (MP 25-26) et un Chiroptère Hipposideros sp, à Delazens.